4 de marzo de 2021 Por Ulrika Beitnere, Segal Lab, UC Davis

Hay un nuevo tipo de nucleasa CRISPR en el bloque de tecnologías genéticas llamado Cas13. ¿Qué tiene de especial y cuál es la principal diferencia con la técnica Cas9 más conocida? Esta es una pregunta que no solo se han hecho muchos padres, ¡sino también científicos! Es posible que desee conocer ¿cómo se puede utilizar Cas13 para el síndrome de Angelman y por qué se debe explorar esa estrategia? “CRISPR nos brinda un nuevo conjunto de herramientas para utilizar en el tratamiento del síndrome de Angelman. El nuevo Cas13 nos permitirá apuntar al antisentido de una manera que puede ser a largo plazo y altamente específica”, afirma el Dr. David Segal, UC Davis.
Primero, veamos su esencia. Tanto las proteínas Cas9 como Cas13 son nucleasas – enzimas que pueden cortar los enlaces fosfodiéster entre los nucleótidos de los ácidos nucleicos. Se ha descubierto que ambas nucleasas son programables en casi cualquier parte del genoma. La principal diferencia es que la nucleasa Cas13 apunta y corta el ARN en lugar del ADN, y esto podría conllevar menos efectos fuera del objetivo marcado. Puesto que, en el síndrome de Angelman, el objetivo es el ARN no codificante largo conocido como transcripción antisentido UBE3A (Ube3a-ATS), resulta óptimo evitar que el ADN sea afectado. El ADN es el guión de cada célula de nuestro cuerpo, mientras que el ARN es la copia de trabajo. Editar la copia de trabajo debería ser más seguro que modificar el original. Con la comprensión de las funciones del ARN y los roles críticos en las enfermedades, así como con el desarrollo de tecnologías relacionadas con el ARN, existe un interés creciente en desarrollar nuevas terapias basadas en ARN, evitando al mismo tiempo al ADN como objetivo.
Recientemente, dos estudios independientes utilizaron Cas9 para silenciar el Ube3a dirigiéndolo a una región del Ube3a-ATS para el síndrome de Angelman en modelos animales. En el primer estudio, Wolter et al. utilizaron un virus adenoasociado (AAV) para administrar Cas9 y descubrieron que Cas9 provocaba que el AAV se insertara en la región diana del Ube3a-ATS en ratones. Solo para refrescar la memoria: el AAV se considera un virus pequeño y seguro que ya se ha utilizado como vehículo de administración para dos terapias genéticas aprobadas por la FDA: Luxturna en 2017 para una distrofia retiniana hereditaria rara y Zolgensma en 2019 para la atrofia muscular espinal. Para Wolter et al., la nucleasa Cas9 hizo que el AAV se insertara en el grupo repetitivo de genes Snord115 ubicado antes del gen Ube3a (Figura 1A). Al apuntar a una gran cantidad de objetivos en ese grupo, pudieron anular el silenciamiento paterno del Ube3a en las neuronas.
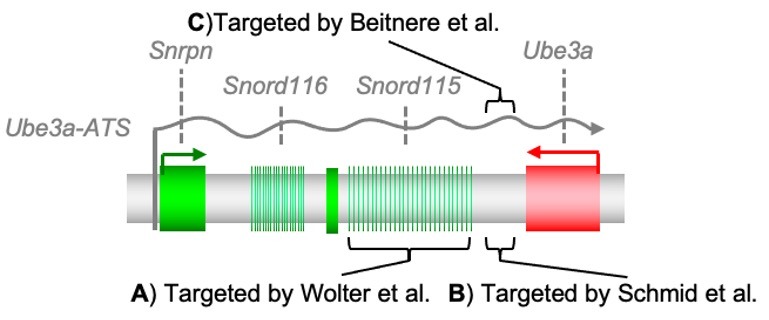
En el segundo estudio, Schmid et al., también usaron AAV para la entrega de Cas9 en una posición diferente del genoma, entre el extremo Ube3a, que se llama 3’ UTR, y el gen Snord115 ( Figura 1B ). A diferencia del estudio de Wolter et al., no observaron mucha inserción de AAV, sino que el Cas9 introdujo inserciones y deleciones cortas (indeles) del ADN en la posición de destino. Estos indeles interfieren con la extensión de Ube3a-ATS y provocan la expresión duradera de la proteína Ube3a del alelo paterno. En resumen, estos dos estudios muestran la prueba de que la nucleasa Cas9 interfiere con el Ube3a-ATS al crear roturas de doble hebra en el ADN en diferentes posiciones y con cantidades variables de integración de AAV, se puede activar la copia paterna del gen Ube3a y rescatar comportamientos del síndrome de Angelman en el modelo de ratón Angelman. En el laboratorio de Segal, estamos trabajando para, en última instancia, hacer el mismo trabajo mientras nos enfocamos en el ARN y no en el ADN, en el que no hay preocupaciones sobre introducir cambios permanentes en el ADN.
Cas13 se está utilizando para apuntar al ARN de Ube3a-ATS sin cambiar el ADN (Figura 1C). Usando AAV para apuntar la nucleasa Cas13 directamente a la forma de ARN de Ube3a-ATS en ratones, también pudimos restaurar la expresión de Ube3a del alelo paterno. Todavía se están realizando varios estudios para que podamos publicar nuestros resultados completos, pero estamos muy entusiasmados con el potencial de Cas13 para lograr esta función. Otros «fármacos dirigidos al ARN» se encuentran en ensayos clínicos, o ya están aprobados, para uso médico: los oligonucleótidos antisentido (ASO) utilizados en los ensayos clínicos para el síndrome de Angelman son un buen ejemplo. Cas13 tiene el potencial de ser un tratamiento ideal para el síndrome de Angelman, al igual que los ASO, pero sin la necesidad de inyecciones intratecales repetidas cada dos meses (Meng et al. 2015). Para otro enfoque dirigido al ARN, la interferencia del ARN, ha sido ya aprobada por la FDA para otros tres tratamientos de enfermedades raras, incluidos Onpattro (2018), Givlaari (2019) y Oxlumo (2020). Cuando se comparó la técnica Cas13 frente a la interferencia directa de ARN, Cas13 ganó en el apartado de precisión, presentando cero fallos en los objetivos (Abudayyeh et al. 2017, Cox et al.2017).
Estudios más interesantes de Cas13 pueden estar avanzando hacia ensayos clínicos. A principios de 2020, cuando el virus COVID-19 se propagaba como la pólvora, un grupo de científicos demostró que Cas13d puede actuar como un tratamiento antiviral al apuntar con éxito a la eliminación del ARN del virus Covid-19 y la gripe A (Abbott et al. 2020). Otro estudio mostró que Cas13a se puede utilizar con éxito para otro trastorno viral: la hepatitis C (Ashraf et al. 2021). El primer estudio que usó Cas13 en animales mostró que Cas13 pudo reparar varios genes en el hígado usando un AAV como vehículo de administración (He et al. 2020).
En resumen, la nueva Cas13 técnica posee la especificidad y la carencia de fallos fuera de objetivos que son ideales para la traducción clínica. Junto con los enfoques emergentes de la eliminación de genes mediada por el ADN de Cas9, una estrategia dirigida al ARN para la interrupción con Cas13 puede ser ventajosa para el tratamiento potencial del síndrome de Angelman y estamos muy emocionados de trabajar en este proyecto.
Referencias
Abbott TR, Dhamdhere G, Liu Y, Lin X, Goudy L, Zeng L, Chemparathy A, Chmura S, Heaton NS, Debs R, Pande T, Endy D, La Russa MF, Lewis DB, Qi LS. Desarrollo de CRISPR como estrategia antiviral para combatir el SARS-CoV-2 y la influenza. Celda. 2020 14 de mayo; 181 (4): 865-876.e12. doi: 10.1016 / j.cell.2020.04.020. Epub 2020 29 de abril. PMID: 32353252; PMCID: PMC7189862.
Abudayyeh OO, Gootenberg JS, Essletzbichler P, Han S, Joung J, Belanto JJ, Verdine V, Cox DBT, Kellner MJ, Regev A, Lander ES, Voytas DF, Ting AY, Zhang F. Selección de ARN con CRISPR-Cas13. Naturaleza. 12 de octubre de 2017; 550 (7675): 280-284. doi: 10.1038 / nature24049. Publicación electrónica del 4 de octubre de 2017. PMID: 28976959; PMCID: PMC5706658.
Cox DBT, Gootenberg JS, Abudayyeh OO, Franklin B, Kellner MJ, Joung J, Zhang F. Edición de ARN con CRISPR-Cas13. Ciencias. 2017 24 de noviembre; 358 (6366): 1019-1027. doi: 10.1126 / science.aaq0180. Publicación electrónica del 25 de octubre de 2017. PMID: 29070703; PMCID: PMC5793859.
Ashraf MU, Salman HM, Khalid MF, Khan MHF, Anwar S, Afzal S, Idrees M, Chaudhary SU. CRISPR-Cas13a mediado por el direccionamiento del posicionamiento de entrada ribosomal interno del virus de la hepatitis C (IRES) como una estrategia antiviral eficaz. Biomed Pharmacother. 19 de enero de 2021; 136: 111239. doi: 10.1016 / j.biopha.2021.111239. Publicación electrónica antes de la impresión. PMID: 33454599.
He B, Peng W, Huang J, Zhang H, Zhou Y, Yang X, Liu J, Li Z, Xu C, Xue M, Yang H, Huang P. Modulación de las funciones metabólicas a través de la eliminación de genes mediada por Cas13d en el hígado. Célula de proteína. 2020 julio; 11 (7): 518-524. doi: 10.1007 / s13238-020-00700-2. PMID: 32185621; PMCID: PMC7095259.
Hebras J, Marty V, Personnaz J, Mercier P, Krogh N, Nielsen H, Aguirrebengoa M, Seitz H, Pradere JP, Guiard BP, Cavaille J. Reevaluación de la participación de Snord115 en la vía del receptor de serotonina 2c en un ratón genéticamente relevante modelo. Elife. 5 de octubre de 2020; 9: e60862. doi: 10.7554 / eLife.60862. PMID: 33016258; PMCID: PMC7673782.
Konermann S, Lotfy P, Brideau NJ, Oki J, Shokhirev MN, Hsu PD. Ingeniería de transcriptomas con efectores CRISPR de tipo VI-D que apuntan a ARN. Celda. 19 de abril de 2018; 173 (3): 665-676.e14. doi: 10.1016 / j.cell.2018.02.033. Publicación electrónica del 15 de marzo de 2018 PMID: 29551272; PMCID: PMC5910255.
Meng L, Ward AJ, Chun S, Bennett CF, Beaudet AL, Rigo F. Hacia una terapia para el síndrome de Angelman dirigiéndose a un ARN largo no codificante. Naturaleza. 19 de febrero de 2015; 518 (7539): 409-12. doi: 10.1038 / nature13975. Publicación electrónica 1 de diciembre de 2014 PMID: 25470045; PMCID: PMC4351819.
